
니코틴아마이드 아데닌 다이뉴클레오타이드 인산

| |
| 식별자 | |
|---|---|
3D 모델 (JSmol)
|
|
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| ECHA InfoCard | 100.000.163 |
| MeSH | NADP |
PubChem CID
|
|
| |
| |
| 성질 | |
| C21H29N7O17P3 | |
| 몰 질량 | 744.416 g·mol−1 |
달리 명시된 경우를 제외하면, 표준상태(25 °C [77 °F], 100 kPa)에서 물질의 정보가 제공됨.
| |
니코틴아마이드 아데닌 다이뉴클레오타이드 인산(영어: nicotinamide adenine dinucleotide phosphate, NADP)은 NADPH를 환원제로 필요로 하는 지질 합성, 핵산 합성과 같은 동화작용에 사용되는 보조 인자이다. 과거에는 트라이포스포피리딘 뉴클레오타이드(영어: triphosphopyridine nucleotide, TPN)로 표기했었다.
NADPH는 환원형, NADP+는 산화형이다. NADP+는 아데닌 잔기와 연결된 리보스의 2' 위치에 추가적인 인산기가 존재한다는 점에서 NAD+와 다르다.
NADP+는 구조뿐만 아니라 기능 또한 NAD+와 비슷한데 일반적으로 탈수소반응의 조효소로 작용한다. 예를 들면 광합성의 명반응에서 물의 광분해로 생성되는 NADPH를 암반응(캘빈 회로)의 1,3-비스포스포글리세르산이 글리세르알데하이드 3-인산으로 환원될 때 NADPH의 전자를 전달해준다.
- NADP+ + 2e- + 2H+ ⇄ NADPH + H+
NADP+/NADPH 쌍으로 생물체 내에서 산화·환원력(redox potential)을 제공한다. 피롤 고리에 탄소-탄소 단일 결합을 이중 결합 구조로 만들어 엽록소 a를 엽록소 b로 만들 때 사용되기도 한다. NADP+(산화형)과 NADPH(환원형)의 2가지 상태를 광합성 생물에서 NADPH는 광합성의 명반응의 전자전달계의 마지막 단계에서 페레독신-NADP+ 환원효소에 의해 생성된다. NADPH는 이산화 탄소를 환원시키기 위해 캘빈 회로의 생합성 반응에 대한 환원력으로 사용된다. NADPH는 이산화 탄소를 포도당으로 전환하는 것을 돕기 위해 사용된다. NADPH는 질소의 순환에서 식물의 동화작용을 위해 질산염을 암모니아로 환원시키는 데에도 필요하다.
동물에서[편집]
동물과 다른 비광합성 생물에서 NADPH의 주요 공급원은 오탄당 인산 경로이다. NADPH를 생성하는 덜 알려진 몇 가지 다른 대사경로가 있으며, 이 대사경로들은 모두 미토콘드리아의 존재에 의존한다. 이러한 과정에서 핵심 효소들은 NADP-결합 말산 탈수소효소(NADP-linked malate dehydrogenase), NADP-결합 아이소시트르산 탈수소효소(NADP-linked isocitrate dehydrogenase), NADP-결합 글루탐산 탈수소효소(NADP-linked glutamate dehydrogenase), 니코틴아마이드 뉴클레오타이드 트랜스하이드로제네이스(nicotinamide nucleotide transhydrogenase)이다.[1] 아이소시트르산 탈수소효소의 메커니즘은 지방세포와 간세포에서 NADPH의 주요 공급원인 것으로 보인다.[2] 또한 미토콘드리아에서 NADH 키네이스(NADH kinase)는 NADH와 ATP를 기질로 사용하여 NADPH와 ADP를 생성한다.
기능[편집]
NADPH는 글루타티온을 재생해서 활성산소의 독성으로부터 보호하는 것을 포함하는 산화환원 반응 및 생합성 반응에 대한 환원당량을 제공한다.[3] NADPH는 콜레스테롤의 합성과 지방산 사슬의 연장과 같은 동화작용에 사용된다.
또한 NADPH 시스템은 면역세포에서 자유 라디칼의 생성을 담당한다. 이러한 라디칼은 호흡폭발이라 불리는 과정에서 병원체를 파괴하는데 사용된다.[4] 이는 방향족 화합물, 스테로이드, 알코올, 약물의 사이토크롬 P450의 하이드록실화에 대한 환원당량의 원천이다.
NADP(H)를 조효소로 사용하는 효소[편집]
- 아드레노독신 환원효소: 이 효소는 대부분의 생물체에 보편적으로 존재한다.[5] 이 효소는 두 개의 전자를 NADPH에서 FAD로 전달한다. 척추동물에서 이 효소는 스테로이드 호르몬을 합성하는 미토콘드리아의 P450 시스템의 사슬에서 첫 번째 효소로 역할을 한다.[6]
- NADP+와 NADPH의 공-막대 모형
-
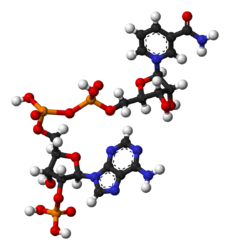 NADP+
NADP+ -
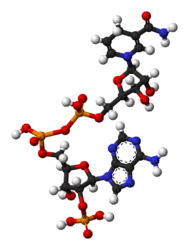 NADPH
NADPH


NADP+의 합성[편집]
NADP+의 기본 골격은 NAD+와 거의 같기 때문에 NAD+의 아데닌 잔기와 연결된 리보스의 2' 위치에 인산기만 붙으면 NADP+가 합성된다.
- NAD+ + ATP → NADP+ + ADP
NADP+의 구조와 특성[편집]
NADP+의 구조는 기본적으로는 NAD+와 거의 같으나(니코틴아마이드와 아데노신으로 구성), 아데노신의 2' 위치에 수소가 아닌 인산기가 붙어있다.
NADP+의 환원 방식은 NAD+의 경우와 같으며 양쪽 모두 뉴클레오타이드를 포함하기 때문에 260 nm 파장에서 흡광도가 가장 크며, 340 nm 파장의 빛을 잘 흡수한다. 그러나 NADP+ 의존성 탈수소효소는 NAD+에는 전혀 활성을 나타내지 않으므로(서로 다른 효소번호[7]를 가지고 있음) NAD+의 방해효과 없이 340 nm에서 NADP+ 의존성 탈수소효소의 활성도를 측정할 수 있다.
NADP+ 및 NADPH의 생리학적 의의[편집]
NADP+/NADPH는 NAD+/NADH와 같은 생체 내의 전자전달 물질이지만 NADH는 미토콘드리아에서 일어나는 산화적 인산화의 전자전달 물질인 반면, NADPH는 엽록체에서 일어나는 광합성의 전자전달 물질이다. 또한 NADPH는 해당과정의 한 종류인 엔트너-듀도로프(Entner-Doudoroff) 경로, 지방산의 합성 및 스테로이드의 합성과정에 사용된다.
NADPH로의 환원[편집]
NADPH를 생성하는 과정은 다음과 같다.
광합성의 명반응에서 광계 I 복합체[편집]
- Fdred + NADP+ → Fdox + NADPH
광계 I 복합체에서 환원된 페레독신(Fd: Feredoxin)이 NADP+로 전자를 전달하여 NADPH가 만들어진다. 이는 비순환적 광인산화에서 일어나며 페레독신-NADP+ 산화환원효소(FNR)가 반응에 관여한다.(순환적 광인산화의 경우, 페레독신에서 플라스토퀴논(PQ: plastoquinone)으로 전자를 전달하여 NADPH가 생성되지 않음) 산소 비발생형 광합성(광합성 세균)에서는 NADP+대신 NAD+가 이용된다.
엔트너-듀도로프 경로[편집]
- 포도당 6-인산 + NADP+ → 6-포스포글루코노-δ-락톤 + NADPH
엔트너-듀도로프 경로와 엠덴-마이어호프-파르나스 경로에서 공통적으로 NAD+가 사용되며, 고균에서만 나타나는 비인산화 엠덴-마이어호프-파르나스 경로에서는 NAD+ 나 NADP+가 사용된다. 아래의 반응에서 NADP+가 사용된다.
- 글리세르알데하이드 + NADP+ → 글리세르산 + NADPH
메탄균의 산화형 불완전 구연산 회로[편집]
NADPH의 산화[편집]
광합성의 캘빈 회로[편집]
- 1,3-비스포스포글리세르산 + NADPH → 글리세르알데하이드 3-인산 + NADP+ + Pi
위의 반응은 글리세르알데하이드 3-인산 탈수소효소가 촉매 역할을 하며, 이 외에 지방산 합성과정에서도 NADP+로 산화된다.
- 아세토아세틸-ACP(ACP: 아실 운반 단백질) + NADPH → β-하이드록시뷰티릴-ACP + NADP+
- 크로토닐-ACP + NADPH → 뷰틸-ACP + NADP+
이 밖에 C4 식물이나 CAM 식물의 광합성에서도 사용된다.
NADP+/NADPH는 주로 동화작용에서 사용되고, NAD+/NADH는 주로 이화작용에서 사용된다.
같이 보기[편집]
각주[편집]
- ↑ Hanukoglu I, Rapoport R (Feb–May 1995). “Routes and regulation of NADPH production in steroidogenic mitochondria”. 《Endocrine Research》 21 (1-2): 231–41. doi:10.3109/07435809509030439. PMID 7588385.
- ↑ Palmer, Michael. “10.4.3 Supply of NADPH for fatty acid synthesis”. 《Metabolism Course Notes》. 6 June 2013에 원본 문서에서 보존된 문서. 6 April 2012에 확인함.
- ↑ Rush GF, Gorski JR, Ripple MG, Sowinski J, Bugelski P, Hewitt WR (May 1985). “Organic hydroperoxide-induced lipid peroxidation and cell death in isolated hepatocytes”. 《Toxicology and Applied Pharmacology》 78 (3): 473–83. doi:10.1016/0041-008X(85)90255-8. PMID 4049396.
- ↑ Ogawa K, Suzuki K, Okutsu M, Yamazaki K, Shinkai S (October 2008). “The association of elevated reactive oxygen species levels from neutrophils with low-grade inflammation in the elderly”. 《Immunity & Ageing》 5: 13. doi:10.1186/1742-4933-5-13. PMC 2582223. PMID 18950479.
- ↑ Hanukoglu I (2017). “Conservation of the Enzyme-Coenzyme Interfaces in FAD and NADP Binding Adrenodoxin Reductase-A Ubiquitous Enzyme”. 《Journal of Molecular Evolution》 85 (5): 205–218. doi:10.1007/s00239-017-9821-9. PMID 29177972.
- ↑ Hanukoglu I (Dec 1992). “Steroidogenic enzymes: structure, function, and role in regulation of steroid hormone biosynthesis”. 《The Journal of Steroid Biochemistry and Molecular Biology》 43 (8): 779–804. doi:10.1016/0960-0760(92)90307-5. PMID 22217824.
- ↑ EC #: Enzyme Commision number